





Особенности строения мышечной клетки. Гипертрофия мышц.
Сегодня поговорим о строении мышечной клетки и о том, как её строение связано с гипертрофией мышц, то есть с увеличением их объёма.
Прежде всего нужно понять общую архитектуру мышцы.
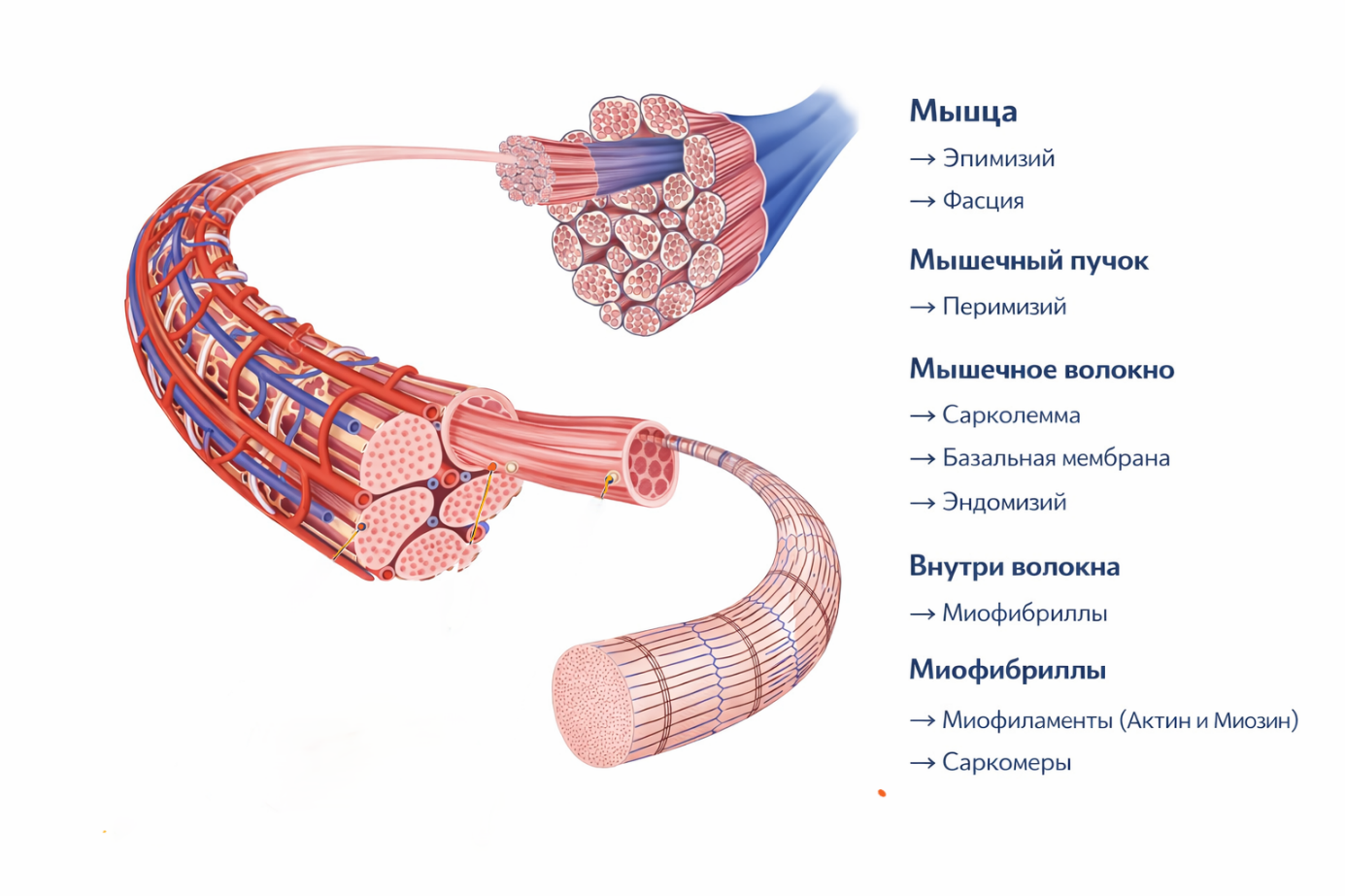
Мышца состоит из мышечных пучков, каждый из которых образован мышечными волокнами. А мышечное волокно — это и есть одна мышечная клетка.
Вся мышца снаружи покрыта соединительнотканной оболочкой — эпимизием. Эпимизий продолжается сухожилием связывающим мышцу с костью. Поверх эпимизия обычно располагается фасция, которая отделяет мышцу от соседних анатомических структур.
Внутри мышцы находятся мышечные пучки, и каждый пучок также окружён соединительнотканной оболочкой — перимизием.
Следующий уровень организации — мышечное волокно. Каждое мышечное волокно представляет собой длинную многоядерную клетку. Его клеточная мембрана называется сарколемма. Снаружи сарколеммы располагается базальная мембрана, а поверх неё находится тонкий слой соединительной ткани — эндомизий.
На этом уровне мышечное волокно густо оплетено капиллярами и нервными окончаниями. Это имеет большое значение, поскольку мышца должна получать достаточное количество кислорода, питательных веществ и нервных сигналов.
Нервные импульсы поступают по мотонейронам и вызывают согласованное сокращение группы мышечных волокон. В этом процессе важную роль играет и внеклеточный матрикс, который механически связывает мышечные волокна между собой и помогает передавать силу сокращения. Эндомизий, перимизий и эпимизий образуют единую соединительнотканную систему мышцы, которая передаёт силу сокращения мышечных волокон на сухожилие.Иннервация мышцы устроена таким образом, чтобы мышечные волокна могли сокращаться синхронно.
Важно отметить, что мышечные клетки значительно отличаются от большинства других клеток организма. Они очень длинные и крупные. Длина мышечного волокна может достигать нескольких сантиметров, поэтому в мышце их относительно немного — счёт идёт скорее на тысячи волокон, а не на миллионы мелких клеток.

Теперь посмотрим, что находится внутри мышечного волокна.
Внутри клетки располагаются многочисленные миофибриллы — длинные белковые структуры, которые проходят вдоль всей клетки. Именно они обеспечивают сокращение мышцы.
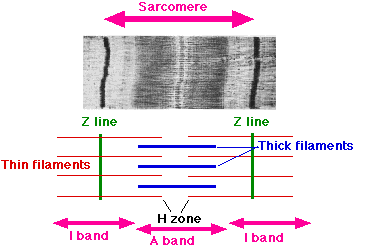
Если рассмотреть миофибриллу под микроскопом, можно увидеть её характерную поперечную исчерченность. Она возникает потому, что миофибриллы состоят из повторяющихся участков — саркомеров.
Саркомер — это элементарная сократительная единица мышцы. Он ограничен с двух сторон структурами, которые называют Z-дисками. Их можно представить как небольшие перегородки внутри мышечного волокна.
К этим Z-дискам прикрепляются актиновые нити — тонкие белковые структуры. Между ними располагаются миозиновые нити — более толстые белковые элементы.
Во время сокращения миозиновые головки связываются с актиновыми нитями и подтягивают их к центру саркомера. Этот процесс часто описывают как систему канатов: миозин словно перехватывает актиновую нить и тянет её на себя.
Поскольку актиновые нити закреплены на Z-дисках, их подтягивание приводит к сближению Z-дисков. Именно это сближение и составляет основу мышечного сокращения.
Процесс происходит симметрично: миозиновые нити тянут актиновые нити с обеих сторон саркомера, поэтому два Z-диска смещаются навстречу друг другу.
Каждая миофибрилла состоит из многочисленных саркомеров
Мышечная клетка (миоцит) значительно отличается от большинства других клеток организма:
- она многоядерная;
- может достигать нескольких сантиметров в длину;
- её цитоплазма (саркоплазма) заполнена миофибриллами;
- миофибриллы состоят из повторяющихся структур — саркомеров;
- эндоплазматический ретикулум представлен специализированной формой — саркоплазматическим ретикулумом, который регулирует уровень кальция.
Если представить это образно, мышечное волокно можно сравнить с гармошкой. Когда гармошку сжимают, её складки сближаются. Примерно такой же принцип работает и в мышце: саркомеры укорачиваются, и всё мышечное волокно становится короче.
Впрчем, в отличиии от гармошки изменение длины одного саркомера очень небольшое, всего 20-30% от длины. Ну и кроме того, в отличии от гармошки усилие прикладывается не снаружи двумя руками, а изнутри.
В одном мышечном волокне находятся тысячи миофибрилл, и все они сокращаются одновременно.
Благодаря этому сокращение происходит синхронно по всей клетке.
Интересно также, что один и тот же Z-диск проходит через всё мышечное волокно, соединяя множество миофибрилл между собой. Это обеспечивает механическую связь всей внутренней структуры клетки и синхронность её работы.
Именно регулярное расположение саркомеров и Z-дисков создаёт характерную поперечную исчерченность мышцы, поэтому скелетную мускулатуру называют поперечно-полосатой.
Поскольку мышечное волокно может достигать нескольких сантиметров в длину, внутри него находится тысячи саркомеров, и именно их согласованная работа приводит к укорочению всей клетки, а затем и всей мышцы.
Гипертрофия мышц
У взрослого человека мышцы растут в основном за счёт гипертрофии, а не за счёт увеличения количества волокон.
То есть:
- не образуются новые мышечные клетки,
- увеличивается диаметр уже существующих волокон.
Рост происходит за счёт увеличения количества миофибрилл и сократительных белков (актина и миозина) внутри клетки.
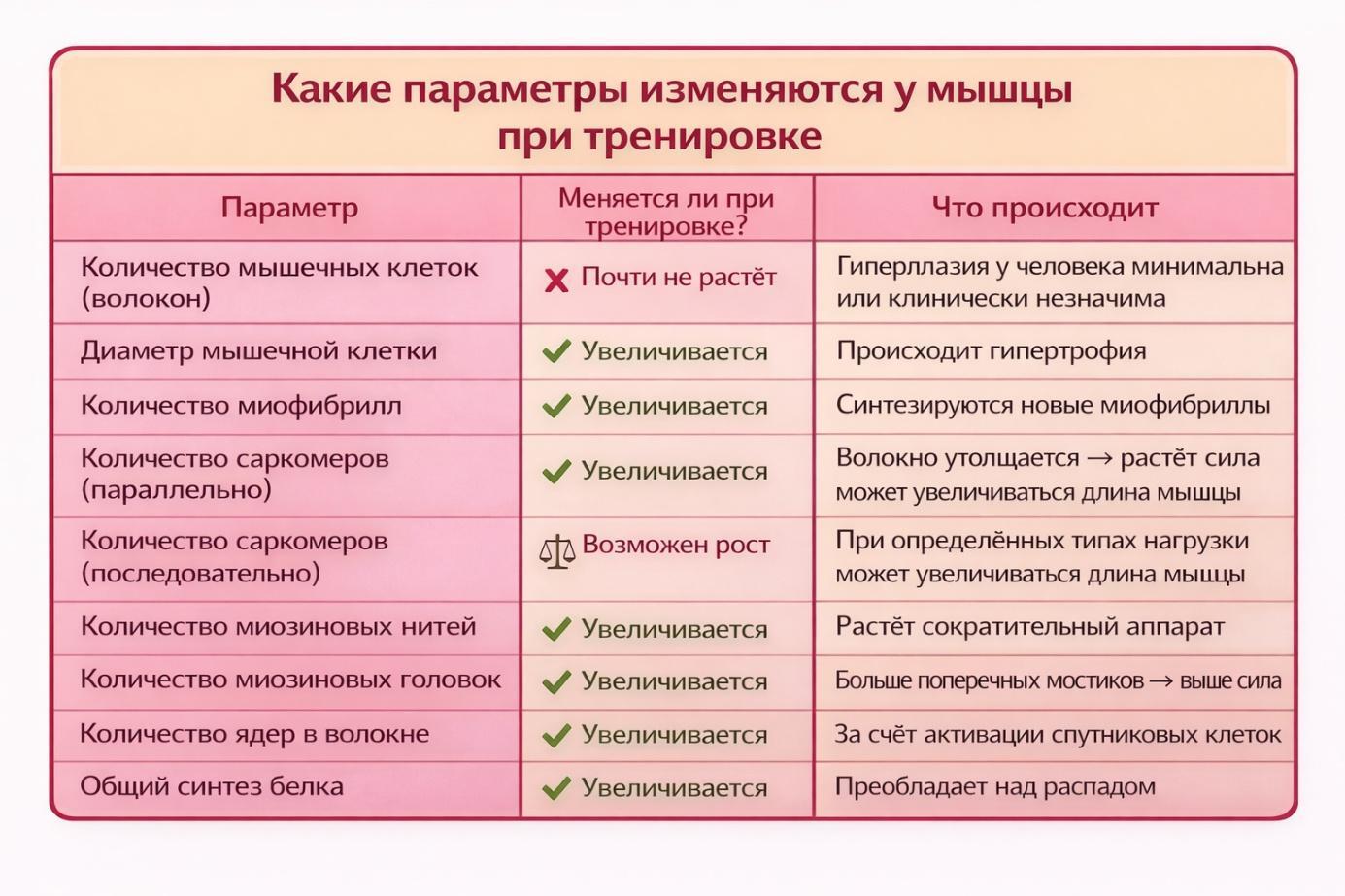
Как правило у нас численность саркомеров увеличивается в толщину, а не в длину. Но есть исключения. Пример нагрузки, увеличивающей длину миофибрилл: эксцентрическая работа мышцы под растягивающей нагрузкой. Например, медленное опускание веса или бег вниз по склону.
Пусковым механизмом роста мышечной массы является механическая нагрузка. Когда мышца работает против значительного сопротивления — например, при силовой работе или подъёме тяжестей — в её структуре возникают процессы, запускающие адаптацию и последующий рост.
Прежде всего механическая нагрузка действует на саркомеры, которые являются основными сократительными единицами мышечного волокна. В саркомерах возникает механическое натяжение, и вся внутренняя архитектура клетки начинает испытывать значительное напряжение. При этом в отдельных участках могут появляться микроповреждения сократительных структур.
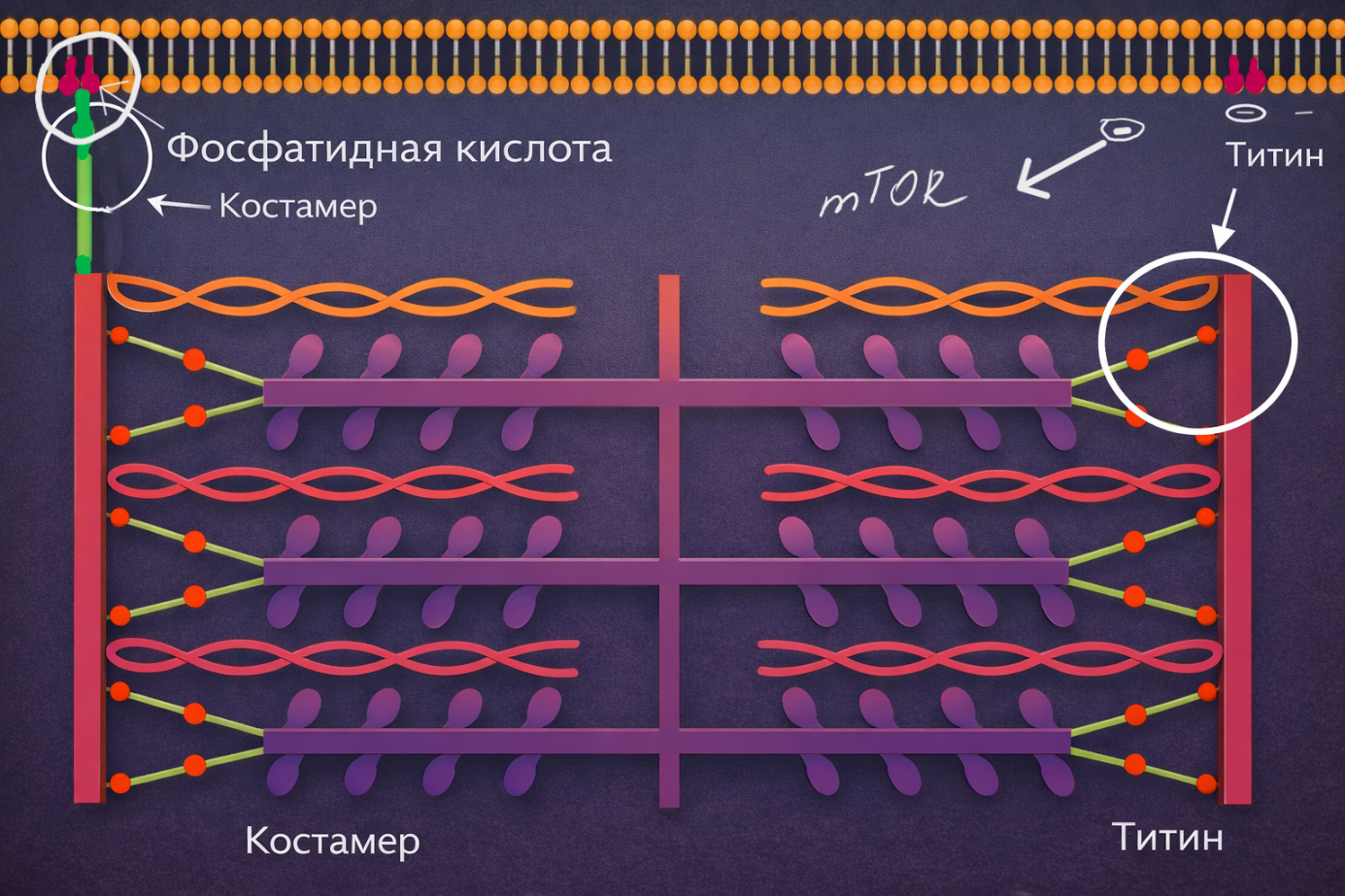
Речь идёт не только о самих актиновых и миозиновых нитях, но и о белках, которые обеспечивают прочность и упорядоченность всей конструкции. Например, важную роль играет белок титин, который соединяет миозиновую нить с Z-диском и выполняет функцию своеобразной молекулярной пружины, стабилизируя саркомер.
Сами Z-диски образованы целым комплексом структурных белков. Эти структуры проходят через всю миофибриллу и выстраиваются в поперечные линии, которые пронизывают всё мышечное волокно.
Далее механическое напряжение передаётся от Z-дисков к периферии клетки. Вблизи сарколеммы Z-диски связаны со специальными белковыми комплексами — костомерами. Костомеры соединяют внутренний сократительный аппарат клетки с её мембраной.
Сарколемма, как и другие клеточные мембраны, представляет собой липидный бислой, в который встроены многочисленные белки. Через систему костомеров механическое напряжение, возникающее внутри саркомеров, передаётся на мембрану клетки и далее — на внеклеточный матрикс мышцы.
Именно в этих зонах передачи силы и возникают важные механические и биохимические сигналы, которые запускают процессы адаптации и роста мышечной ткани.
Одним из таких сигналов является образование фосфатидной кислоты. Это особый мембранный фосфолипид, который играет роль важной сигнальной молекулы и участвует в запуске процессов гипертрофии мышечной клетки.
При механической нагрузке, когда напряжение передаётся через саркомеры, Z-диски и систему костомеров к сарколемме, происходит активация ряда ферментов, прежде всего фосфолипазы D. Этот фермент действует на компоненты мембраны и приводит к образованию фосфатидной кислоты непосредственно в липидном бислое сарколеммы.
Повышение концентрации фосфатидной кислоты является важным сигналом для клетки. Она взаимодействует с рядом внутриклеточных белков и активирует один из ключевых регуляторов клеточного роста — сигнальный комплекс mTOR (mechanistic Target Of Rapamycin).

Активация этого сигнального пути запускает каскад реакций, которые приводят к усилению синтеза белка в мышечной клетке. В результате увеличивается образование сократительных белков — прежде всего актина и миозина, а также других структурных компонентов саркомеров.
Одновременно усиливается синтез белков цитоскелета, ферментов энергетического обмена и элементов саркоплазматического ретикулума. Всё это приводит к увеличению количества миофибрилл внутри мышечного волокна и, как следствие, к увеличению его диаметра.
Таким образом механическая нагрузка через систему саркомеров, Z-дисков и костомеров передаётся на клеточную мембрану, где запускаются биохимические сигналы роста, одним из ключевых элементов которых является образование фосфатидной кислоты и последующая активация сигнального пути mTOR. Именно этот механизм лежит в основе гипертрофии мышечной ткани.
Активация сигнального пути mTOR приводит не только к усилению синтеза белков за счёт собственного генетического аппарата мышечной клетки. Одновременно запускаются процессы, которые активируют так называемые сателлитные клетки — мышечные стволовые клетки, расположенные между сарколеммой и базальной мембраной мышечного волокна.
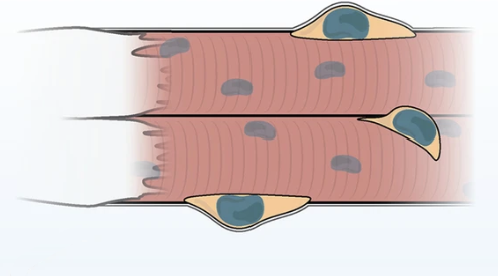
Сателлитные клетки начинают делиться, после чего часть из них дифференцируется и сливается с существующим мышечным волокном. При этом они передают мышечной клетке свои ядра, а вместе с ними и дополнительный генетический материал.
Это важно потому, что у мышечного волокна существует определённое ограничение, которое называют лимитом ядерного домена. Каждое ядро способно обслуживать только определённый объём цитоплазмы и обеспечивать синтез ограниченного количества белков. Пока этот предел не превышен, мышечная клетка может увеличиваться за счёт собственного генетического аппарата. Однако по мере роста волокна возникает необходимость в дополнительных ядрах, которые смогут поддерживать дальнейший синтез белков.
Именно эту функцию и выполняют сателлитные клетки. Сливаясь с мышечным волокном, они увеличивают количество ядер в клетке, тем самым расширяя её возможности по синтезу белков и поддержанию структуры сократительного аппарата.
Таким образом, гипертрофия мышечного волокна обеспечивается двумя взаимосвязанными механизмами: с одной стороны, активацией внутриклеточных сигнальных путей роста, таких как mTOR, усиливающих синтез белков, а с другой — увеличением числа ядер за счёт слияния сателлитных клеток. Благодаря этому мышечное волокно получает возможность продолжать увеличиваться в размере и усиливать свою сократительную способность.
Механическая нагрузка
↓
деформация костомеров и сарколеммы
↓
активация фосфолипазы D
↓
образование фосфатидной кислоты в мембране
↓
активация mTOR
↓
усиление синтеза белка
↓
выделение факторов роста
↓
активация сателлитных клеток
Спутниковые клетки.
В ответ на механическую нагрузку или повреждение мышцы эти клетки активируются и начинают делиться. Обычно их деление носит асимметричный характер: одна клетка остаётся сателлитной, сохраняя запас стволовых клеток, а другая превращается в миобласт. Миобласт затем дифференцируется и сливается с существующим мышечным волокном, добавляя ему новое ядро. Миобласт в свою очередт может делиться дальше, но уже симметрично. Таким образом увеличивается количество ядер и расширяются возможности клетки по синтезу сократительных белков, что обеспечивает рост мышечного волокна и его адаптацию к нагрузке.
С возрастом активность сателлитных клеток постепенно снижается. Это связано с несколькими факторами: ограниченным числом делений клеток (правило Хейфлика), укорочением теломер, а также изменениями гормонального фона — снижением уровня тестостерона, соматотропного гормона и инсулиноподобного фактора роста (IGF-1). В результате способность мышечной ткани к регенерации и увеличению числа ядер уменьшается, что способствует постепенному развитию саркопении — возрастного снижения мышечной массы и силы
На столько укорачивается саркомер при сокращении
20–30%
Миофибрилл содержит одно мышечное волокно
1000+
Снижают активность сателлитных клеток с возрастом: теломеры, гормоны, лимит делений
3 фактора
Другие материалы

Получить первичную оценку документов
Пришлите ваши выписки/снимки/анализы и мы вышлем вам первичную оценку ваших документов