





Физиология мышечного сокращения
Физиологические механизмы сокращения скелетной мускулатуры.
Сегодня мы поговорим о строении скелетных мышц и механизмах их сокращенияТакже мы коснёмся роли таких макроэлементов, как кальций, натрий, калий, магний и фосфор.
То есть речь пойдёт о пяти из семи основных макроэлементов организма человека. Макроэлементами называют химические элементы, которые необходимы нам в сравнительно больших количествах — их суточная потребность измеряется сотнями миллиграммов или граммами.

Если говорить о распространённости дефицитов, то ситуация неодинакова.
Недостаточное потребление магния, кальция и калия встречается достаточно часто, особенно при несбалансированном рационе, бедном овощами, фруктами, цельными продуктами и источниками белка.
Дефицит фосфора и натрия при обычном питании развивается крайне редко, поскольку они широко представлены в пищевых продуктах и хорошо регулируются организмом.
Отдельно стоит упомянуть кальций. Его недостаточное потребление чаще наблюдается у подростков и у женщин старшего возраста.
У подростков это связано с интенсивным ростом костной ткани: потребность в кальции в этот период повышена, и при несоответствии рациона потребностям может формироваться дефицит по питанию.
У женщин после менопаузы снижение уровня эстрогенов приводит к усилению резорбции костной ткани. Это не означает, что кальция становится мало в крови — его уровень поддерживается довольно строго. Речь идёт о постепенной потере кальция из костей и снижении их минеральной плотности, что повышает риск остеопороза.
Таким образом, роль макроэлементов в организме тесно связана как с питанием, так и с гормональной регуляцией и возрастными изменениями.
Роль этих макроэлементов в работе скелетной мускулатуры трудно переоценить. Без них невозможны ни передача нервного импульса, ни запуск сокращения, ни сам механический акт работы мышцы, ни её последующее расслабление.
И сегодня мы разберёмся, почему каждый из них — кальций, натрий, калий, магний и фосфор — является обязательным звеном в этой сложной и точно настроенной системе.
Начало сокращения: команда из мозга
Любое движение начинается с сигнала из головного мозга.
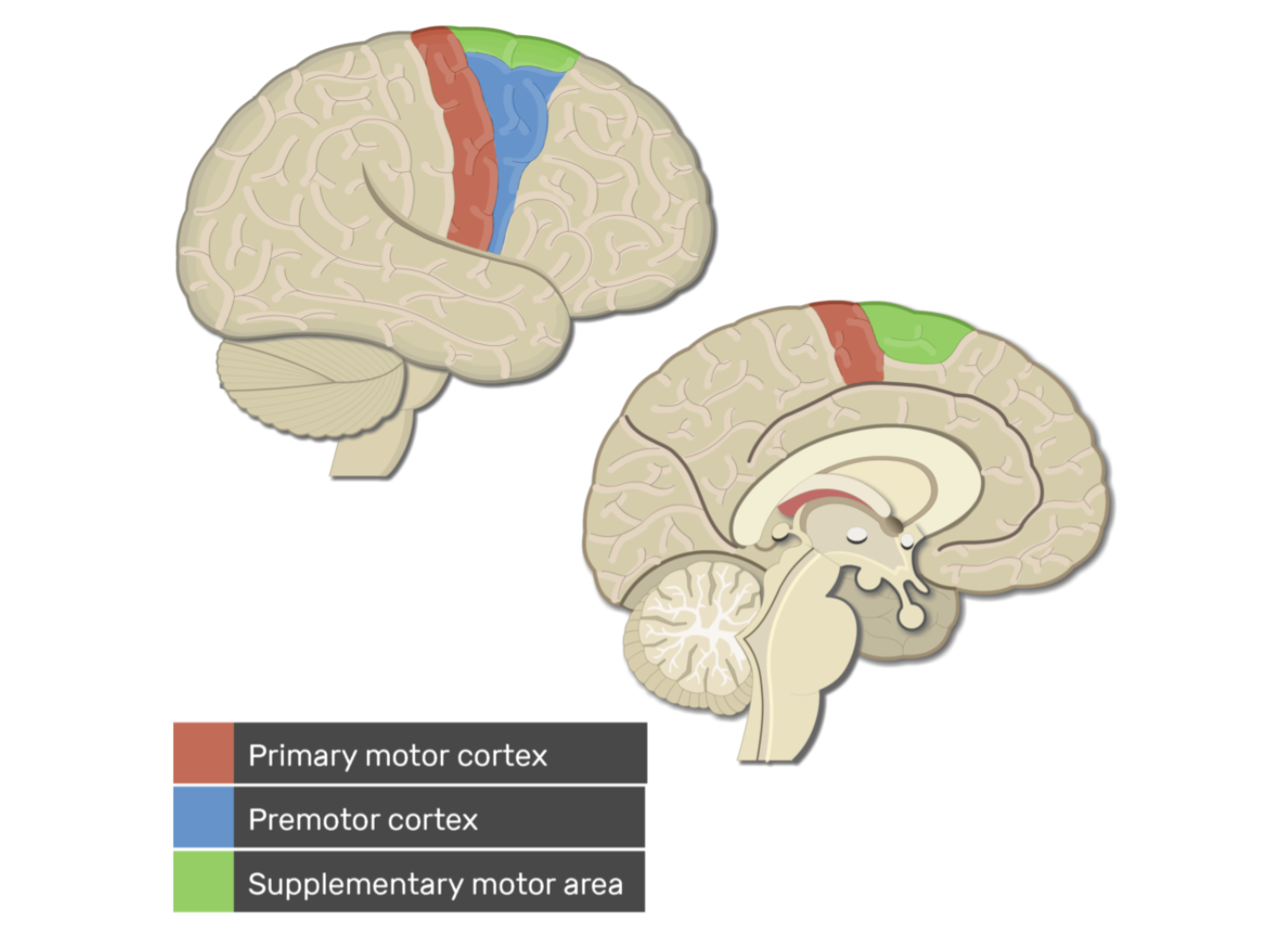
Supplementary motor area (SMA) — дополнительная моторная область
(зелёная область) участвует в планировании сложных движений
Premotor cortex — премоторная кора (синяя область)отвечает за подготовку движения
Primary motor cortex — первичная моторная кора (красная область) главный источник двигательных команд. Она расположена в прецентральной извилине. Здесь находятся крупные пирамидные нейроны (клетки Беца), аксоны которых формируют: кортико-спинномозговой (пирамидный) путь. Именно эти нейроны отправляют сигнал: мозг → спинной мозг → мотонейрон → мышца.
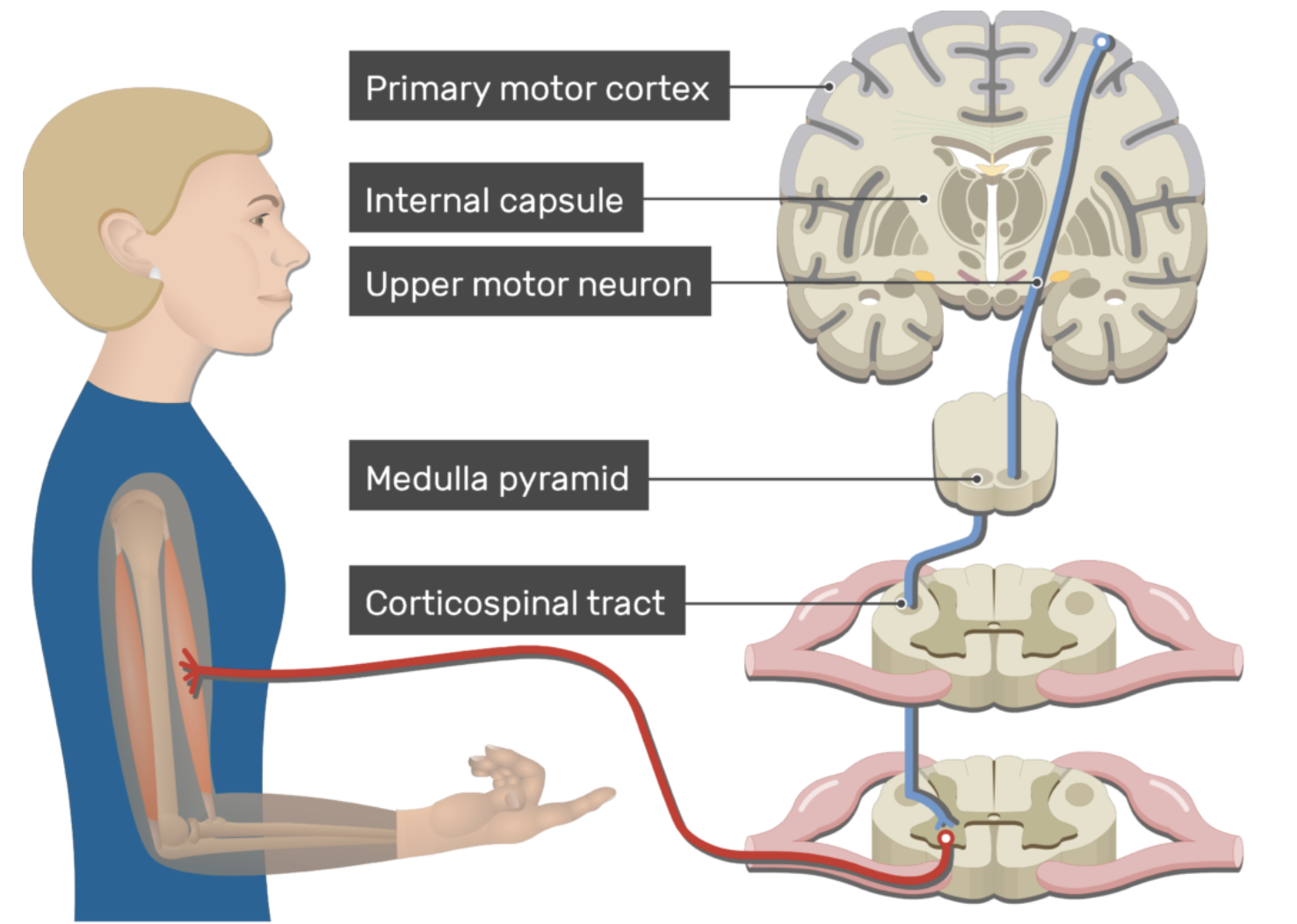
От коры головного мозга до мышцы сигнал передаётся через два нейрона. Первый расположен в моторной коре и называется верхним (центральным) мотонейроном. Его аксон образует кортико-спинномозговой, или пирамидный, путь и проводит импульс вниз. В продолговатом мозге большая часть этих волокон перекрещивается, поэтому правое полушарие управляет мышцами левой половины тела и наоборот. Далее сигнал поступает ко второму нейрону — нижнему (периферическому) мотонейрону, который находится в передних рогах спинного мозга и посылает аксон к мышце. Для мышц головы и лица путь короче: верхний мотонейрон из коры соединяется с ядрами черепных нервов в стволе мозга (в мосту и продолговатом мозге), где и расположены периферические мотонейроны, управляющие мимическими мышцами.
Нервный импульс распространяется по мотонейрону в виде волны возбуждения.
Как распространяется нервный импульс
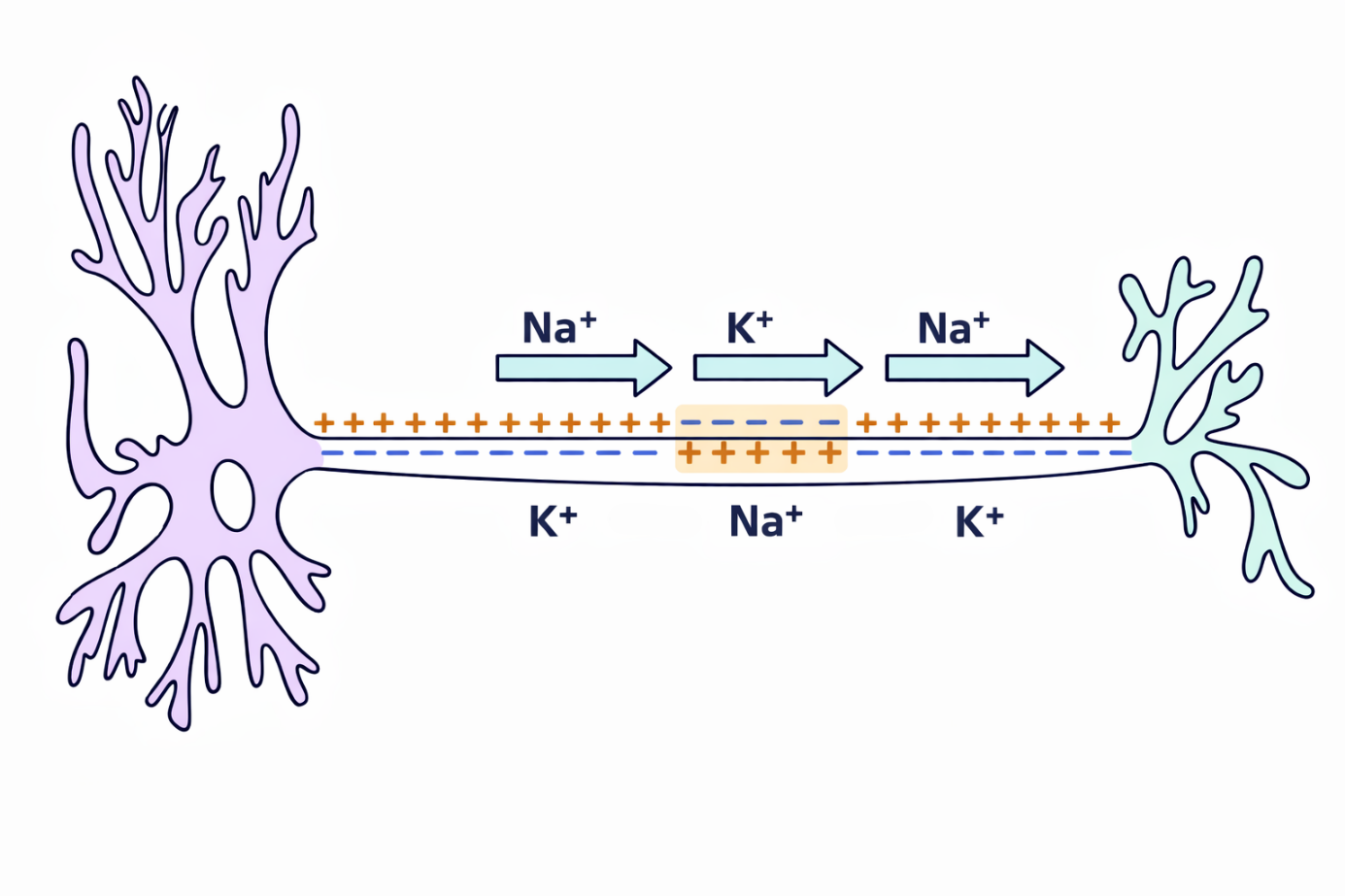
Теперь важно понять, как именно распространяется нервный сигнал.
Импульс передаётся от тела нейрона по его длинному отростку — аксону. Скорость распространения сигнала может превышать 100 метров в секунду, особенно в миелинизированных нервных волокнах.
Это происходит благодаря явлению, которое называется потенциал действия. Он возникает в мембране нервной клетки и связан с движением ионов натрия и калия через специальные белковые структуры мембраны — ионные каналы.
Здесь важно не путать каналы и насосы. Это разные механизмы.
Ионные каналы пропускают ионы пассивно, по градиенту концентрации.
Ионные насосы, наоборот, переносят ионы активно, затрачивая энергию АТФ.
В состоянии покоя мембрана нейрона имеет электрический потенциал примерно –70…–90 милливольт. Это состояние поддерживается прежде всего работой натрий-калиевого насоса, который выкачивает натрий из клетки и закачивает калий внутрь.
Когда нейрон получает возбуждение и достигается пороговый уровень примерно –55 милливольт, открываются натриевые каналы. Натрий начинает быстро входить в клетку, и мембрана деполяризуется — её потенциал становится менее отрицательным.
Затем открываются калиевые каналы. Калий начинает выходить из клетки, и мембрана постепенно реполяризуется, то есть возвращается к отрицательному потенциалу.
Однако калиевые каналы работают немного дольше, чем нужно, поэтому возникает небольшое переразряжение мембраны — так называемая гиперполяризация. Потенциал становится даже немного более отрицательным, чем в состоянии покоя.
После этого в работу снова вступает натрий-калиевый насос, который постепенно восстанавливает исходное распределение ионов и возвращает мембрану к нормальному потенциалу покоя.
Таким образом, последовательность процессов выглядит так:
пороговое возбуждение → открытие натриевых каналов → деполяризация → открытие калиевых каналов → реполяризация → кратковременная гиперполяризация → восстановление потенциала покоя с помощью натрий-калиевого насоса.
Именно эта волна изменений электрического потенциала и распространяется вдоль мембраны аксона, передавая нервный сигнал по всей длине нервного волокна.
Передача импульса на мышечное волокно
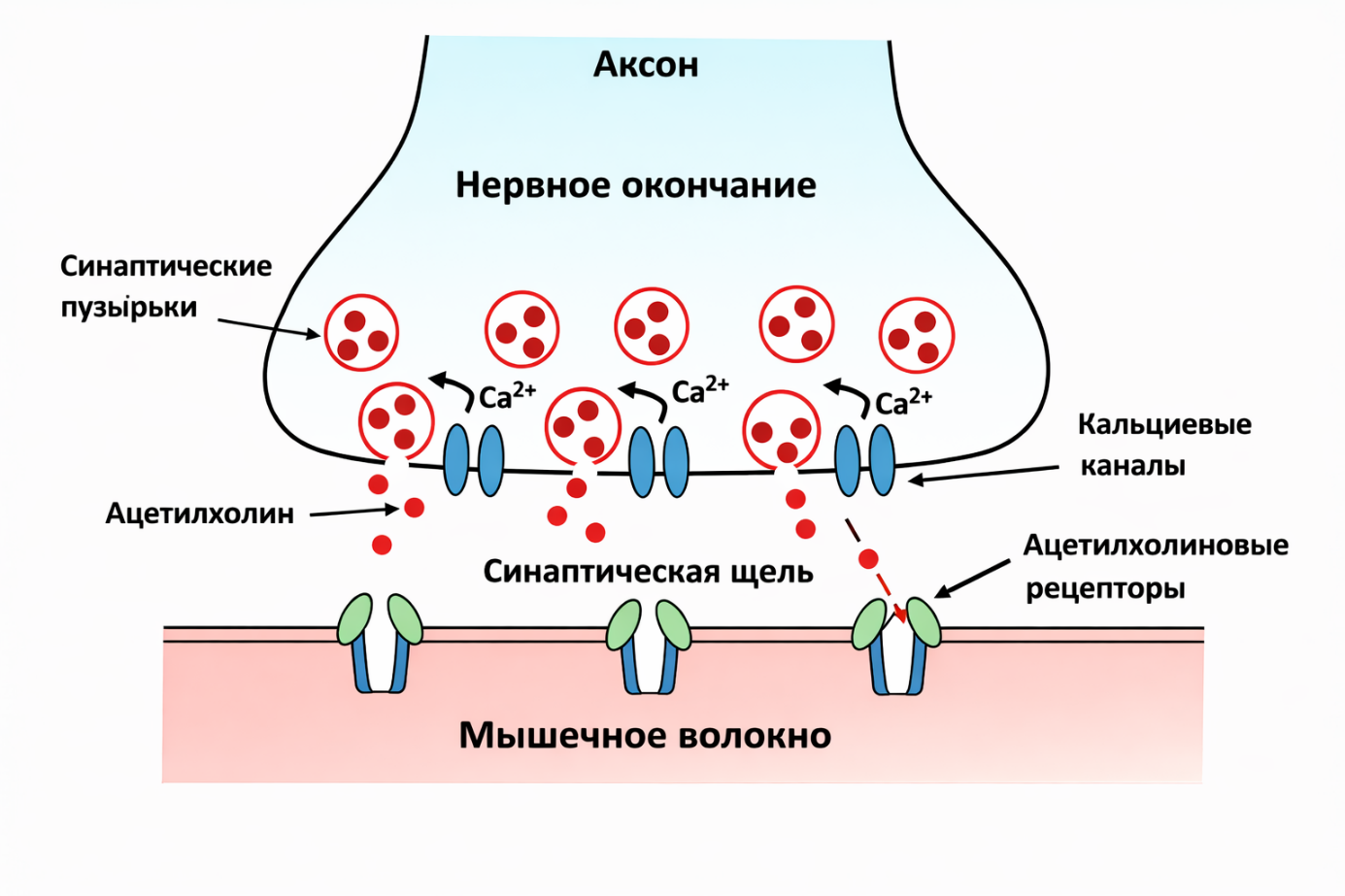
Ацетилхолин связывается с рецепторами на мембране мышечной клетки и запускает дальнейшую передачу сигнала, которая в конечном итоге приводит к сокращению мышцы.
Когда потенциал действия достигает пресинаптического окончания:
- Мембрана деполяризуется.
- Открываются кальциевые каналы.
- Концентрация Ca²⁺ локально резко возрастает.
И вот здесь главный момент:
Именно повышение внутриклеточного кальция запускает экзоцитоз.
Ca²⁺ связывается с белком синаптотагмином на мембране синаптических пузырьков.
Далее:
- активируется комплекс SNARE-белков,
- пузырёк сливается с мембраной,
- ацетилхолин (или другой медиатор) выбрасывается в синаптическую щель.
Ацетилхолин связывается с никотиновыми ацетилхолиновыми рецепторами (nAChR) на постсинаптической мембране мышечного волокна (в области моторной пластинки).
Эти рецепторы являются лигандзависимыми катионными каналами. После связывания ацетилхолина канал открывается и становится проницаемым для положительно заряженных ионов — прежде всего для Na⁺ и K⁺.
Движение ионов через открытый канал происходит по их электрохимическому градиенту, то есть под действием разницы концентраций и электрического потенциала по обе стороны мембраны.
Поскольку в покое:
- концентрация Na⁺ значительно выше снаружи клетки,
- внутри клетки поддерживается отрицательный мембранный потенциал,
натрий преимущественно входит в клетку.
Калий, напротив, имеет высокую внутриклеточную концентрацию и может выходить наружу, однако его выход выражен слабее, чем вход натрия.
В результате суммарный ток направлен внутрь клетки, что приводит к деполяризации постсинаптической мембраны и формированию концевого пластинчатого потенциала.
Если деполяризация достигает порогового значения, запускается потенциал действия мышечного волокна и начинается процесс сокращения.
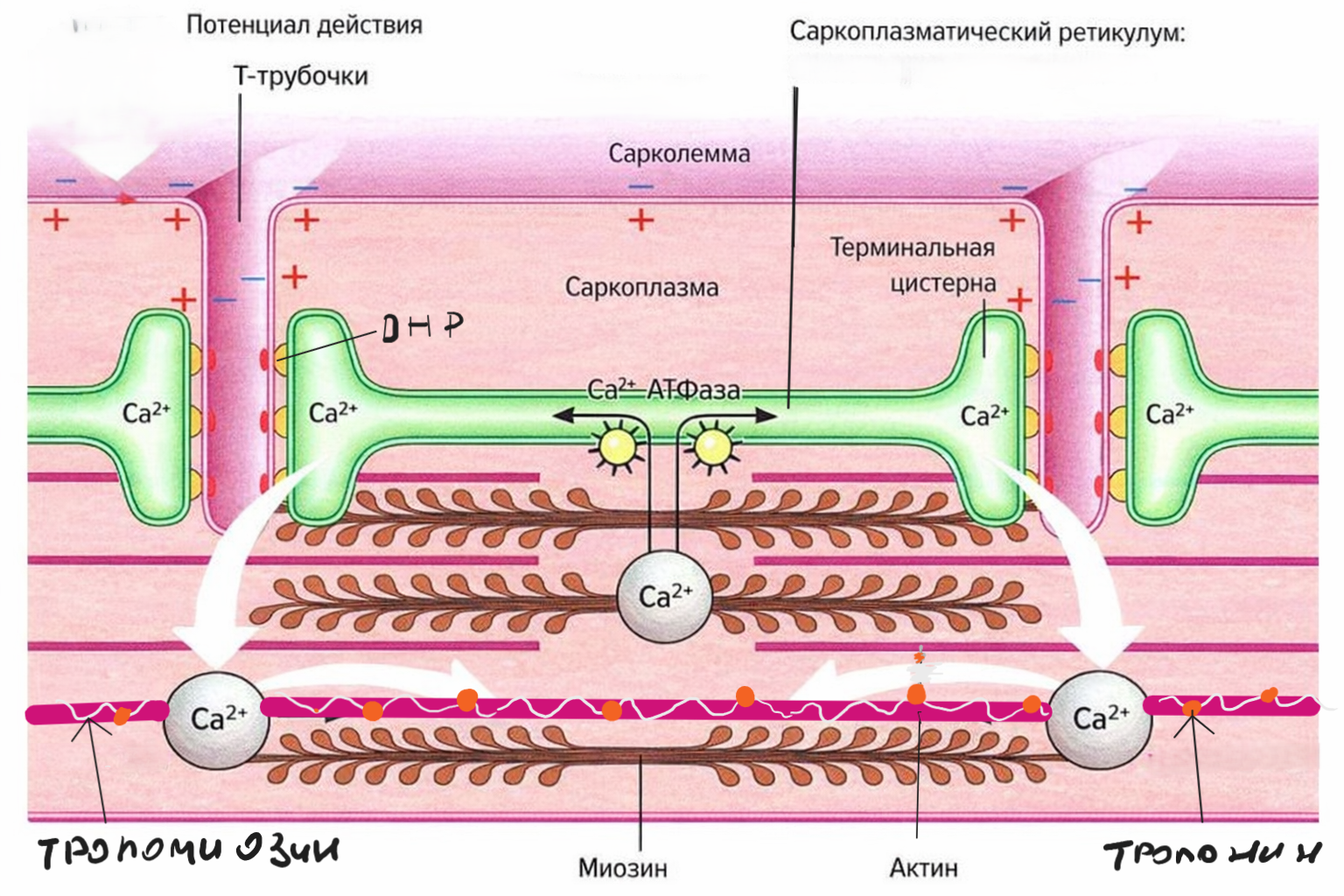
Распространение сигнала внутри мышечной клетки
Потенциал действия распространяется по всей сарколемме и уходит вглубь клетки через Т-трубочки.
В мембране Т-трубочек находятся DHP-рецепторы, которые механически связаны с рианодиновыми рецепторами саркоплазматического ретикулума.
В результате этой активации из саркоплазматического ретикулума в цитоплазму выбрасывается кальций.
Роль кальция в сокращении
Именно кальций запускает процесс сокращения.
Он связывается с тропонином, что приводит к смещению тропомиозина и открытию активных центров актина.
После этого миозиновые головки начинают взаимодействовать с актиновыми нитями — происходит скольжение нитей относительно друг друга, и саркомер укорачивается.
Так сокращаются миллионы саркомеров одновременно, и мышечное волокно укорачивается как единое целое.
Расслабление мышцы
Для расслабления кальций должен быть удалён из цитоплазмы.
Саркоплазматический ретикулум с помощью кальциевых насосов (Ca²⁺-АТФазы) закачивает кальций обратно внутрь.
Когда мы говорим о сокращении мышцы, чаще всего вспоминаем кальций. Он действительно запускает процесс, связываясь с тропонином и открывая доступ миозину к актину. Но механический цикл невозможен без фосфора и магния.
Фосфор входит в состав молекулы АТФ — аденозинтрифосфата. В этой молекуле три фосфатные группы, и энергия запасена не просто «в фосфоре», а в связях между этими фосфатами. Когда миозиновая головка связывает АТФ, она расщепляет его до АДФ и неорганического фосфата. Именно в этот момент миозиновая головка переходит в так называемое «взведённое» состояние — она накапливает энергию.
После этого миозин связывается с актином. Затем происходит высвобождение фосфата, и именно этот шаг запускает силовой ход — миозиновая головка меняет форму и тянет актиновую нить. Это не «передача фосфора» актину, а изменение конфигурации самого миозина, вызванное предыдущим расщеплением АТФ. Так химическая энергия превращается в механическое движение.
В клетке АТФ практически никогда не существует сам по себе. Он связан с ионом магния. Поэтому правильнее говорить не просто «АТФ», а комплекс Mg–АТФ. Именно в таком виде молекула распознаётся миозиновой АТФазой и может быть расщеплена. Без магния этот процесс протекал бы крайне неэффективно. По сути, магний обеспечивает правильную работу энергетического механизма сокращения.
Но его роль этим не ограничивается.
Магний влияет и на регуляцию кальция. Он частично конкурирует с кальцием за участки связывания в белках, стабилизирует мембраны, снижает чрезмерную возбудимость. В состоянии покоя часть участков тропонина занята магнием. Когда концентрация кальция повышается, кальций вытесняет магний в регуляторных зонах и запускает сокращение.
Однако расслабление мышцы происходит не потому, что магний «включает релаксацию», а потому что кальций удаляется из цитоплазмы обратно в саркоплазматический ретикулум. И здесь магний снова необходим — он участвует в работе кальциевых насосов, которые возвращают Ca²⁺ внутрь ретикулума.
Каждый цикл работы насоса переносит два иона Ca²⁺ внутрь ретикулума на одну молекулу АТФ. Именно поэтому расслабление мышцы — это тоже энергозависимый процесс. Если АТФ нет, насос останавливается, кальций остаётся в цитоплазме, и мышца не может расслабиться (пример — трупное окоченение).
Скорость распространения нервного импульса по миелинизированному волокну
100 м/с
Потенциал покоя мембраны нейрона — стартовая точка любого движения
–70 мВ
Перекачивает насос за один цикл, возвращая мышцу в состояние покоя
2 иона Ca²⁺
Другие материалы

Получить первичную оценку документов
Пришлите ваши выписки/снимки/анализы и мы вышлем вам первичную оценку ваших документов